
Peptide library screening is one of those techniques that looks deceptively simple on paper but rewards the researchers who understand its real mechanics. At its core, what is peptide library screening? It's a systematic method for identifying peptide sequences that bind to or modulate a biological target by exposing large, diverse peptide collections to selection pressure in iterative cycles. The field formally calls this affinity selection from combinatorial peptide libraries, and it sits at the intersection of molecular biology, combinatorial chemistry, and bioinformatics. Done well, it accelerates hit identification by orders of magnitude compared to one-at-a-time synthesis and testing.
Table of Contents
- Key takeaways
- What peptide libraries are and how peptide library screening works
- Major library screening technologies and platforms
- Applications in drug discovery and translational research
- Challenges and practical tips for better screening outcomes
- My take on where peptide library screening is heading
- How Innovabiotech can support your screening program
- FAQ
Key takeaways
| Point | Details |
|---|---|
| Screening is iterative | Each round of binding, washing, and elution enriches the highest-affinity peptides from millions of candidates. |
| Platform choice matters | Phage display, mRNA display, and yeast display differ in library size, throughput, and selection conditions. |
| Controls prevent false positives | Counter-selection and naïve library sequencing distinguish true binders from nonspecific artifacts. |
| Applications extend beyond binding | Peptide libraries support epitope mapping, T-cell antigen discovery, immunopeptidomics, and functional cell-based assays. |
| Bioinformatics is non-negotiable | Deep sequencing and computational analysis convert raw enrichment data into ranked, validated hits. |
What peptide libraries are and how peptide library screening works
Understanding peptide libraries starts with scale. A peptide library is a large, diverse collection of peptide sequences, typically ranging from thousands to trillions of unique molecules, each differing in amino acid composition, length, or chemical modification. These collections are synthesized chemically or expressed biologically, and the goal is always the same: give your target as many candidates as possible so that rare, high-affinity binders can be found.
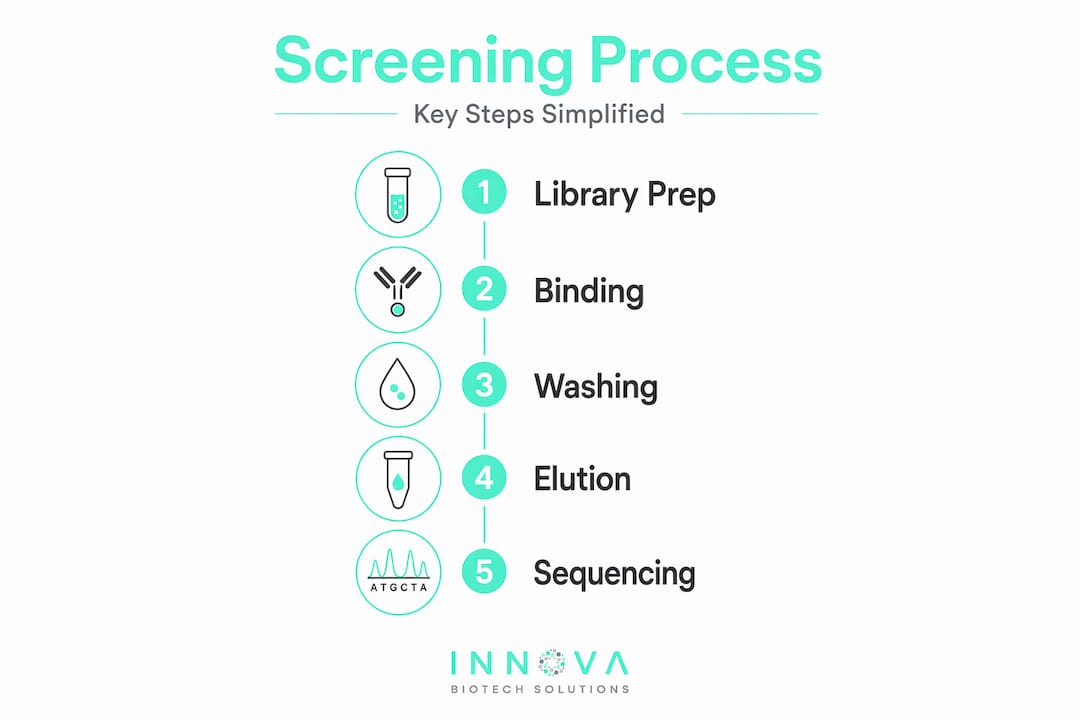
The screening process itself works through iterative cycles of binding, washing, and elution. You expose the library to an immobilized or solution-phase target, remove non-binders and weak binders by washing, then elute the peptides that stayed attached. Those survivors go into the next round, where selection pressure typically increases.
Here is what that cycle looks like in practice:
- Binding: The peptide library contacts the target under defined conditions of concentration, pH, and temperature.
- Washing: Stringency is adjusted to remove nonspecific binders, often by increasing salt concentration or detergent.
- Elution: Bound peptides are released by competitive displacement, pH shift, or protease digestion.
- Amplification or recovery: In display formats, eluted clones are amplified before the next round; in synthetic libraries, eluted peptides go directly to sequencing.
- Sequencing and ranking: After two to five rounds, deep sequencing ranks enriched sequences by frequency and fold-enrichment to identify top candidates.
Pro Tip: Run at least one round of sequencing on the naïve, pre-selection library. You need that baseline to separate genuine enrichment from sequences that were simply overrepresented at the start.
The power of this approach is that you are not guessing which peptide to test. You are letting the target itself pick its best binders from an enormous population.
Major library screening technologies and platforms
No single platform dominates peptide library screening. The right choice depends on your library size requirements, whether your target tolerates cellular contexts, and how harsh a selection environment you need.
| Platform | Library size | Selection conditions | Key advantage | Key limitation |
|---|---|---|---|---|
| Phage display | ~10^10 | Mild, aqueous | Genotype-phenotype linkage; easy amplification | Propagation bias; limited to display-compatible peptides |
| mRNA display | ~10^15 | Harsh conditions tolerated | Largest diversity; covalent mRNA-peptide link | Inefficient puromycin ligation can reduce yield |
| Yeast display | ~10^8 | Cell-compatible | FACS-based quantitative sorting | Smaller library size; expression constraints |
Phage display and biopanning
Phage display remains the most widely deployed library screening technique in drug discovery. Peptides are expressed as fusions on the surface of bacteriophage, and each phage particle carries both the peptide and the gene encoding it. This genotype-phenotype linkage is what makes the system work. The biopanning workflow involves immobilizing the target, incubating with the phage library, washing away non-binders with progressively increasing stringency, eluting bound phage, and then re-amplifying in bacteria before the next round.

After three to five rounds, the pool collapses to a handful of dominant sequences. Sequencing those sequences reveals your hits.
mRNA display for harsh selections
When your selection conditions demand elevated salt, organic solvents, or extreme pH ranges that would kill cells or degrade viral particles, mRNA display is the right tool. The peptide is covalently linked to its encoding mRNA via a puromycin moiety at the 3' end. After translation, the puromycin inserts into the ribosome and forms a stable mRNA-peptide covalent fusion. This means even after harsh washing, each surviving molecule retains its genetic barcode.
Library sizes reach approximately 10^15 unique sequences, far exceeding what phage display achieves. The tradeoff is that inefficient puromycin ligation reduces the fraction of molecules that form valid fusions, which lowers the effective diversity that participates in selection.
Yeast display and FACS-based sorting
Yeast display takes a fundamentally different approach to peptide profiling. Peptides are expressed as fusions on the yeast cell surface, and the entire selection happens through fluorescence-activated cell sorting. Combining yeast surface display with flow cytometry lets you gate simultaneously on display level and binding signal, which directly addresses one of the biggest problems in peptide screening: separating high-affinity binders from peptides that appear to bind only because they are overexpressed.
Pro Tip: In yeast display campaigns, gate on display expression first, then apply the binding gate. Normalizing binding signal to display level prevents low-displaying, high-signal clones from contaminating your selection.
Applications in drug discovery and translational research
Peptide library screening generates hits across a remarkably wide set of biological problems. The most direct application is identifying therapeutic peptide candidates that modulate enzymes, receptors, and protein-protein interactions. But the technique's reach has grown well beyond that.
The broad application landscape now includes:
- Epitope mapping: Overlapping peptide libraries tile across an antigen sequence to identify the precise residues recognized by antibodies or T-cell receptors. This is standard practice in vaccine development and immunotherapy design.
- Diagnostics: Short peptides that bind biomarkers with high specificity can be identified from libraries faster and more cheaply than antibody-based discovery, making this an appealing route for diagnostics programs.
- Immunopeptidomics: Large user-defined peptide libraries improve targeted mass spectrometry performance dramatically. One 2026 study showed that matching library composition to the peptide universe enabled recovery of more than 75% of expected sequences per injection, with detection sensitivity down to 0.1 fmol in complex biological backgrounds.
- T-cell antigen discovery: Functional screening now goes well beyond binding. A 2026 study used SCT peptide libraries with multi-parameter profiling to identify 2,188 antigen-specific CD4+ T cells across 22 participants from a 23-element library, demonstrating that cell-based functional screens can extract population-level immune data from relatively compact libraries.
The shift toward functional and cellular screening is one of the most significant developments in the field. You are no longer just asking "does this peptide bind?" You are asking "does this peptide do something relevant inside a physiological context?"
Challenges and practical tips for better screening outcomes
Even well-designed screening campaigns produce misleading results if the controls and workflow decisions are not right. These are the most common failure points and how to address them.
-
Run counter-selections. Before exposing your library to the intended target, pan it against the capture surface alone, or against an unrelated protein bound to that surface. Peptides that bind the streptavidin, resin, or plate material rather than your target will otherwise dominate later rounds.
-
Monitor enrichment round-by-round with sequencing. Do not wait until the final round to sequence. Tracking enrichment dynamics across rounds tells you whether the selection is collapsing too fast, suggesting amplification bias, or whether diversity is being maintained appropriately through mid-campaign.
-
Watch for phage propagation bias. Some phage clones replicate faster than others regardless of binding affinity. Propagation-driven library collapse can fill your final pool with fast growers rather than true binders. Cross-referencing enrichment with control selections without target is the practical fix.
-
Increase stringency progressively. Do not jump to high-stringency washing in round one. Start mild and tighten with each subsequent round. You preserve early-round diversity while still converging on the best binders by the end.
-
Validate hits orthogonally. Sequencing enrichment is a guide, not a final answer. Confirm your top candidates by resynthesizing as discrete peptides and testing affinity using surface plasmon resonance, isothermal titration calorimetry, or a cell-based functional assay relevant to your program.
Pro Tip: For any display-based campaign, sequence the input library before selection. This lets you calculate enrichment fold over starting frequency, which is far more informative than absolute read counts alone.
My take on where peptide library screening is heading
I've spent a significant amount of time working through peptide screening workflows across different target classes, and one pattern stands out consistently: researchers underestimate how much of their screening success or failure is determined before the first selection round runs. Library design, target preparation, and control architecture matter more than platform choice in most cases.
What I find genuinely exciting right now is the integration of next-generation sequencing into every round, not just the final one. Watching how a library collapses in real time completely changes how you interpret results. You stop asking "what did I get at the end?" and start asking "what dynamics shaped that outcome?" Those are very different questions, and the second one leads to much better experimental decisions.
I've also come to believe that no single display technology gives you the complete picture. In programs where resources allow, running phage display and yeast display in parallel on the same target is not redundant. The two methods apply different selection pressures, and the intersection of their hit lists tends to be far more reliable than either list alone.
The biggest misconception I encounter is that more rounds always means better results. In reality, over-selection is a serious problem. The library collapses, propagation biases dominate, and you end up with confident-looking sequences that don't validate. Stopping at three to four rounds and sequencing deeply is almost always the better strategy.
The future I see involves tighter coupling between physical peptide library campaigns and computational prediction, using screening data as training sets for models that predict binding from sequence. That feedback loop will compress discovery timelines considerably.
— Hooman
How Innovabiotech can support your screening program
If the complexity of peptide library screening has you rethinking your current workflow, that's a reasonable reaction. Designing libraries, selecting platforms, and interpreting enrichment data correctly takes expertise that most research teams are still building.
Innovabiotech offers custom peptide design services built specifically for drug discovery researchers who need libraries tailored to their target class and selection format. The team also provides virtual screening services that complement physical screening by computationally pre-filtering candidates, reducing the library complexity you need to synthesize or display. For programs moving from hit identification into lead optimization, protein engineering support is available to refine peptide structure-activity relationships with precision. Whether you are launching a first screening campaign or troubleshooting an existing one, Innovabiotech works alongside your team through every stage.
FAQ
What is peptide library screening in simple terms?
Peptide library screening is a method for finding peptide sequences that bind to or modulate a biological target by testing large collections of peptides in iterative selection cycles. Each round enriches higher-affinity binders until top candidates can be identified through sequencing.
How does phage display differ from mRNA display?
Phage display links peptide identity to a bacteriophage particle and uses biological amplification between rounds, while mRNA display creates a covalent mRNA-peptide bond enabling library sizes up to 10^15 and selections under harsher conditions that would destroy phage.
What are the main benefits of peptide screening over traditional synthesis?
Peptide screening evaluates millions to trillions of sequences simultaneously, identifying hits in weeks rather than the years it would take through sequential synthesis and testing. It also uncovers unexpected binding motifs that rational design would not predict.
How do you avoid false positives in peptide library screening?
Running counter-selections against the capture surface, sequencing the naïve input library, and tracking enrichment dynamics across all rounds are the core strategies. Controlling for nonspecific binders is as important as the positive selection itself.
What role does next-generation sequencing play in modern screening?
NGS has transformed peptide library analysis by enabling round-by-round monitoring of enrichment dynamics, detection of propagation biases, and quantitative affinity ranking across thousands of sequences simultaneously rather than picking and Sanger-sequencing individual clones.