
TL;DR:
- Effective linker design connects protein domains while preserving function and folding within fusion proteins.
- Using computational modeling like geometry models and structure prediction tools improves the accuracy of linker selection and reduces trial-and-error.
Linker design for chimeric proteins is defined as the rational selection and engineering of peptide sequences that connect two or more protein domains within a single fusion construct. These sequences, typically 5 to 50 amino acids in length, act as dynamic molecular hinges that control domain spacing, orientation, and relative mobility. The wrong linker collapses a functional construct. The right one preserves each domain's activity while enabling the chimeric protein to behave as a coherent unit. Innovabiotech works with researchers at this exact decision point, applying computational modeling and target-specific analysis to get linker selection right the first time.
What are the key linker properties in chimeric proteins?
The three core linker categories are flexible, rigid, and semi-flexible. Each serves a distinct structural purpose, and choosing the wrong type is one of the most common reasons a chimeric construct fails to perform as expected.

Flexible linkers use glycine-serine motifs, most commonly (GGGGS)n repeats tested at one to four repeat units. Glycine provides rotational freedom, and serine adds solubility. These linkers work well when the two domains need spatial freedom to fold independently or when the exact distance between domains is uncertain. The trade-off is reduced structural predictability, which can complicate crystallography and binding assays.
Rigid linkers use alpha-helical sequences such as (EAAAK)n. They maintain a fixed distance between domains and prevent unwanted interdomain interactions. Rigid linkers suit applications where domain proximity would cause steric interference or where one domain must present a binding surface in a defined orientation.
Semi-flexible linkers fall between these two extremes. They combine short stretches of glycine-serine with proline or alanine residues to allow limited movement while maintaining partial directionality. This category has grown in use as researchers recognize that most real-world fusion proteins need controlled, not unlimited, flexibility.
| Linker type | Typical motif | Best application |
|---|---|---|
| Flexible | (GGGGS)1–4 | Independent domain folding, uncertain spacing |
| Rigid | (EAAAK)n | Fixed orientation, preventing domain crosstalk |
| Semi-flexible | Mixed Gly/Ser/Pro | Partial mobility with directional control |
| Cleavable | Protease recognition site | Conditional domain separation in vivo |
Beyond type, linker length and composition together determine whether each domain reaches its binding partner or folds correctly. A linker that is too short forces domains into steric conflict. One that is too long introduces unstructured regions that reduce expression yield and increase proteolytic vulnerability.
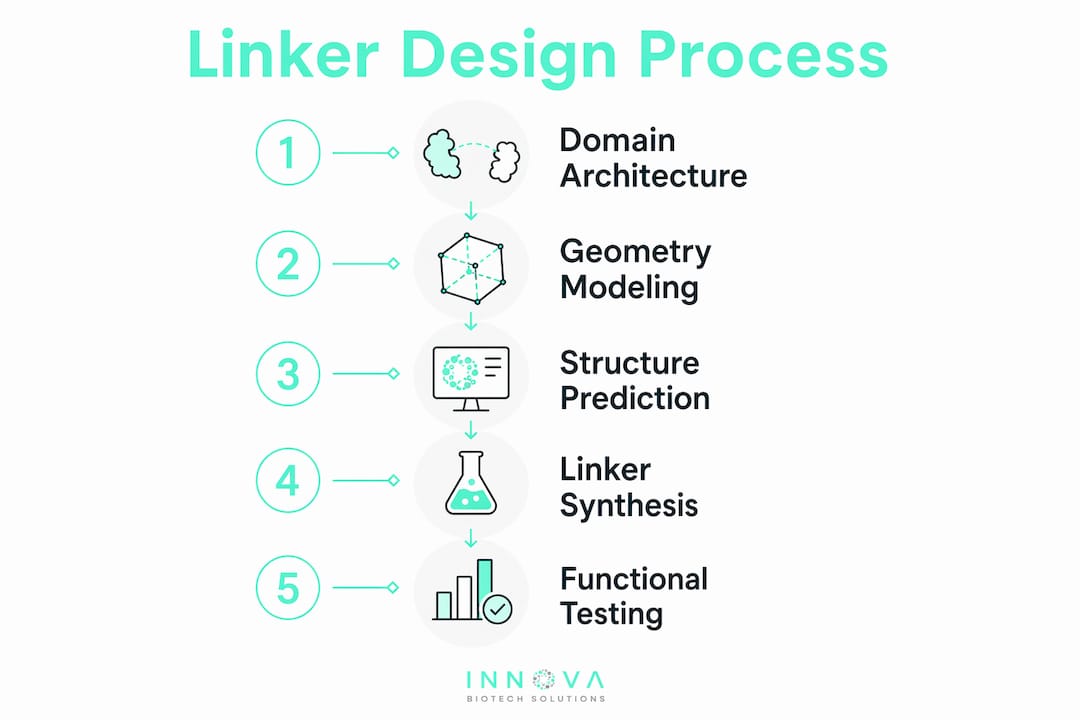
How do you design linkers optimized for a target chimeric protein?
Rational linker design starts with the target protein's domain architecture, not with a standard sequence pulled from a database. Domain fusion orientation and the order in which domains are arranged critically influence folding and functional integrity. N-terminal fusions behave differently from C-terminal fusions, and swapping the order of two domains can shift activity by an order of magnitude.
Use physical geometry models first
Modeling linkers as worm-like chains or helical rods predicts domain reachability and steric clashes before you synthesize anything. The worm-like chain model treats the linker as a flexible polymer and calculates the probability that the two domain termini stay within a functional distance. The helical rod model applies when you expect the linker to adopt a defined secondary structure. Both approaches replace empirical trial-and-error with quantitative predictions. That shift alone reduces the number of constructs you need to test in the wet lab.
Integrate structure prediction tools
AlphaFold2 has become a standard checkpoint in linker evaluation. You build the full chimeric sequence, run a structure prediction, and measure RMSD between the predicted structure and the isolated domain structures. A low RMSD confirms the linker preserves each domain's fold. A high RMSD signals that the linker is distorting one or both domains, and you revise before synthesis.
Multimodal deep learning frameworks that combine sequence representations with Cα-Cα distance matrix-derived substructure features go further. These models predict functional linker performance more accurately than classical flexibility classification alone. They enable systematic customization of linkers for specific fusion proteins rather than relying on category-level heuristics.
Pro Tip: Before running any structure prediction, strip your construct of purification tags. A ten-histidine tag placed near the linker region can distort predicted geometry and mislead your RMSD analysis.
| Design step | Tool or method | Output |
|---|---|---|
| Domain geometry analysis | Worm-like chain / helical rod model | Reachability probability, steric risk |
| Structure prediction | AlphaFold2 | RMSD, domain fold preservation |
| Sequence-structure integration | Multimodal deep learning | Functional linker ranking |
| Experimental validation | Fluorescence, binding assay, crystallography | Confirmed activity |
Researchers working on de novo protein design will recognize this workflow. The same principles that govern de novo fold design apply directly to linker selection within chimeric constructs.
What are common challenges in linker design and how do you fix them?
The most persistent problem in linker design is treating it as an afterthought. Researchers finalize both domains, then select a generic (GGGGS)3 linker because it worked in a previous construct. That logic fails because linker performance is target-protein dependent. A linker that works perfectly for one domain pair can cause misfolding or loss of function in another.
Domain misfolding and steric clashes
Domain misfolding occurs when the linker forces a domain terminus into a conformation that conflicts with the domain's natural folding pathway. Steric clashes happen when the linker is too short and the two domains physically collide. Both problems show up as reduced expression yield, aggregation, or loss of binding activity. The fix is to model the geometry before synthesis and test at least two linker lengths bracketing your predicted optimum.
Loss of function after tag addition
Purification tags such as ten-histidine sequences alter crystallization behavior and can interfere with linker function. Tag removal improved crystal morphology and crystallization rates in documented cases. This is not a minor detail. If your construct includes a His-tag adjacent to the linker, test the tag-free version before concluding the linker itself is the problem.
Pro Tip: Run a parallel expression test with and without your purification tag. If yield or activity improves after tag removal, the tag was the variable, not the linker sequence.
- Screen at least three linker lengths for each new domain pair
- Test both N-terminal and C-terminal fusion orientations before committing to one
- Use computational peptide screening to rank candidates before wet-lab synthesis
- Validate with a functional assay specific to the domain of interest, not just SDS-PAGE
Systematic screening beats intuition every time. The goal is to reduce the experimental space with computational predictions, then confirm with targeted wet-lab tests.
Which linker designs have proven effective in recent applications?
Evidence from recent computational and experimental studies points to clear patterns in which linker types succeed for specific domain classes.
For BMP2 chimeric proteins, flexible (GGGGS)1–3 linkers at the N-terminus produce lower RMSD values than rigid (EAAAK)n linkers. Lower RMSD means the predicted structure aligns more closely with the native domain structure. That result confirms the linker preserves structural integrity rather than distorting the domain fold. For collagen-binding chimeras specifically, N-terminal flexible linkers outperform their rigid counterparts on this metric.
For DARPin fusions, the pattern reverses. Short semi-flexible and rigid linkers suit DARPin domains, while flexible linkers suit UBA domains. DARPins are repeat proteins with a defined, elongated structure. They need a linker that maintains orientation rather than one that introduces rotational freedom. UBA domains, which bind ubiquitin through a surface-exposed helix, benefit from the spatial freedom that flexible linkers provide.
Deep learning-based linker design has produced a third category of results. Optimized linkers designed by multimodal deep learning outperform rigid controls at approximately 85% activity in sfGFP fluorescence assays. That benchmark matters because sfGFP fluorescence is a direct readout of correct protein folding. A linker that preserves sfGFP fluorescence at 85% of the unlinked control has maintained the domain's functional fold through the fusion.
| Application | Best linker type | Key outcome |
|---|---|---|
| BMP2 collagen-binding chimera | Flexible (GGGGS)1–3, N-terminal | Lower RMSD, preserved domain fold |
| DARPin fusion | Semi-flexible or rigid | Maintained orientation, functional binding |
| UBA domain fusion | Flexible | Spatial freedom for surface-exposed binding |
| sfGFP fluorescence assay | Deep learning-optimized | ~85% activity vs. rigid control |
Domain selection and linker choice are inseparable decisions. The evidence above shows that no single linker type dominates across applications. The domain's structural class, its binding mechanism, and the fusion orientation all determine which linker category to start with.
Key Takeaways
Effective linker design requires matching linker type and length to the specific structural and functional demands of each domain pair, not applying a universal sequence.
| Point | Details |
|---|---|
| Linker type is domain-specific | Flexible linkers suit UBA domains; rigid or semi-flexible linkers suit DARPins. |
| Physical modeling reduces wet-lab waste | Worm-like chain and helical rod models predict reachability before synthesis. |
| Purification tags affect linker performance | Remove His-tags adjacent to linkers before concluding the linker sequence is the problem. |
| Deep learning outperforms classical design | Multimodal frameworks combining sequence and structural features achieve ~85% activity in functional assays. |
| Domain order matters as much as linker sequence | N-terminal vs. C-terminal fusion orientation changes folding outcomes independently of linker choice. |
Why I think most researchers underestimate linker context
After working through a significant number of chimeric protein design projects, the pattern I keep seeing is the same: researchers spend months optimizing each domain individually, then treat the linker as a connector they can pick in an afternoon. That framing is wrong, and it costs time.
The linker is not passive. It participates in the folding pathway of both adjacent domains. A rigid linker placed between two domains that need to reorient during binding will suppress function even if both domains are perfectly designed in isolation. I have seen constructs with excellent individual domain activity fail entirely because the linker locked them into a geometry that prevented the conformational change needed for binding.
The shift I advocate for is treating linker design as a first-class design problem, not a post-hoc connection step. That means running geometry models before you finalize domain boundaries, testing fusion orientation as a variable, and using structure prediction to screen linker candidates computationally before committing to synthesis. The protein engineering for therapeutics field is moving in this direction, and the researchers who adopt this workflow early will spend less time troubleshooting failed constructs.
The other thing I would emphasize is the purification tag problem. It is mentioned in the literature but rarely treated as a design variable in practice. If your tag sits between the linker and a domain terminus, it changes the effective linker length and geometry. That is a confounding variable in every experiment you run until you account for it. Remove the tag, retest, and then decide whether the linker needs to change.
AI-driven design tools are making this entire process faster. But the underlying logic, match linker geometry to domain requirements, has not changed. The tools just let you test more candidates in silico before touching a pipette.
— Hooman
Innovabiotech's approach to chimeric protein and linker design
Innovabiotech brings together computational modeling and experimental design expertise to support researchers working on complex fusion protein projects.
The team at Innovabiotech applies structure-based linker selection, physical geometry modeling, and deep learning-informed screening to each chimeric protein design project. For researchers who need custom linker sequences validated against specific domain targets, Innovabiotech's peptide design services cover the full workflow from sequence generation through bioinformatics validation. Every project is scoped to the specific domain pair, fusion orientation, and functional assay requirements of the client's research goals. Contact Innovabiotech to discuss your construct's linker requirements directly with the design team.
FAQ
What is linker design for chimeric proteins?
Linker design for chimeric proteins is the process of selecting and engineering peptide sequences that connect protein domains within a fusion construct to preserve function and structural integrity. Linkers typically range from 5 to 50 amino acids and are chosen based on the target domain's structural class and fusion orientation.
What are the best linkers for chimeric proteins?
The best linker depends on the target protein. Flexible (GGGGS)n linkers suit domains that need spatial freedom, while rigid (EAAAK)n or semi-flexible linkers suit domains requiring fixed orientation, such as DARPins.
How does linker length affect chimeric protein stability?
Linker length directly affects domain spacing and folding. A linker that is too short causes steric clashes between domains, while one that is too long introduces unstructured regions that reduce expression yield and increase proteolytic sensitivity.
Can computational tools predict linker performance?
Yes. Tools such as AlphaFold2 and multimodal deep learning frameworks that combine sequence and structural features predict linker performance before wet-lab synthesis, reducing the number of constructs that need experimental testing.
Should purification tags be included in linker design planning?
Yes. Purification tags placed adjacent to the linker region change the effective linker geometry and can interfere with crystallization and binding assays. Testing the tag-free construct is a standard troubleshooting step before revising the linker sequence itself.